Кросинговер: повний розбір генетичної рекомбінації в мейозі
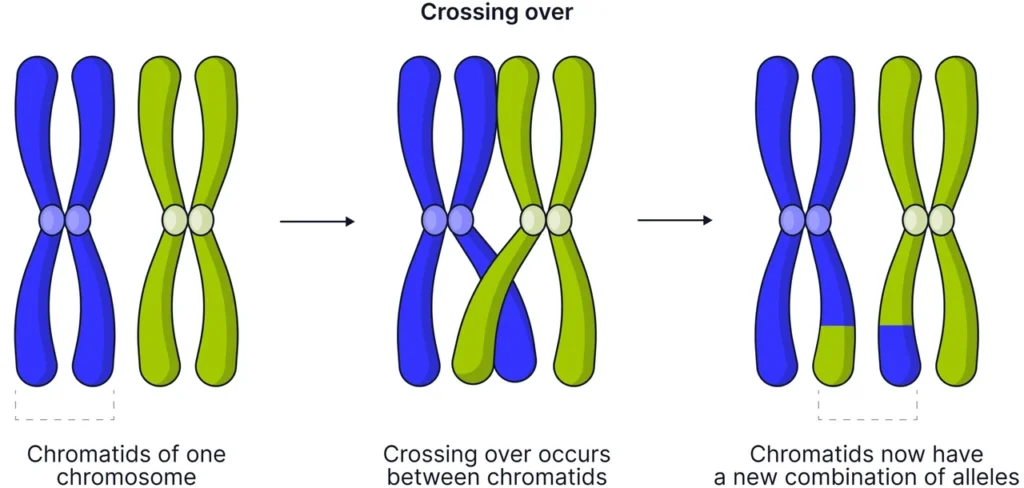
Кросинговер являє собою фундаментальний процес у біології, під час якого гомологічні хромосоми обмінюються генетичним матеріалом у профазі першого поділу мейозу. Цей обмін створює нові комбінації алелів, забезпечуючи унікальність кожного нащадка та рушійну силу еволюції. Без нього статеве розмноження втратило б свою головну перевагу — неймовірну генетичну різноманітність, яка дозволяє популяціям адаптуватися до змін навколишнього середовища протягом тисячоліть.
На молекулярному рівні кросинговер починається з контрольованих розривів подвійного ланцюга ДНК, ініційованих білком Spo11, і завершується формуванням хіазм, які не лише перетасовують гени, а й гарантують правильне розходження хромосом під час анафази. Сучасні дослідження розкривають складну мережу білків, які регулюють цей процес, роблячи його одним із найточніших механізмів у клітині, де навіть найменша помилка може призвести до серйозних наслідків для потомства.
Історія вивчення кросинговеру, що почалася з робіт Томаса Моргана понад століття тому, перетворила генетику з описової науки на точну дисципліну, де частота рекомбінації стала інструментом для побудови карт геномів і розуміння того, як спадковість працює на рівні хромосом.
Де і коли відбувається кросинговер
Кросинговер розгортається виключно в мейозі — спеціалізованому поділі, що зменшує кількість хромосом удвічі для утворення гаплоїдних гамет. Він локалізується в профазі першого мейотичного поділу, точніше на стадії пахітени, коли гомологічні хромосоми вже повністю спарені.
Спочатку в лептотені хромосоми конденсуються, стаючи видимими як тонкі нитки. У зиготені відбувається кон’югація — точне попарне зближення гомологів по всій довжині, утворюючи біваленти. Саме тоді формується синаптонемальний комплекс — білкова «блискавка», яка утримує пари хромосом разом і створює ідеальні умови для обміну. На стадії пахітени, коли біваленти виглядають як товсті нитки, і відбувається власне кросинговер: несестринські хроматиди розриваються та переплітаються.
У диплотені синаптонемальний комплекс починає розпадатися, але хромосоми залишаються з’єднаними в точках перехрестя — хіазмах. Ці видимі структури є прямим наслідком кросинговеру. Нарешті в діакінезі хромосоми максимально конденсуються, ядерна оболонка руйнується, і біваленти вишиковуються для метафази I. Кожна хіазма — це не просто слід обміну, а й механічний якір, без якого гомологи можуть розійтися неправильно.
Історія відкриття кросинговеру
У 1909 році бельгійський цитолог Франс Янссенс описав хіазми — перехрестя хромосом у мейозі земноводних. Він припустив, що ці структури пов’язані з обміном матеріалом, але не мав генетичних доказів. Прорив стався в 1911 році, коли американський генетик Томас Хант Морган разом зі студентом Альфредом Стёртевантом вивчав мутації в X-хромосомі плодової мушки Drosophila melanogaster.
Аналізуючи результати схрещувань, Морган помітив, що деякі комбінації ознак з’являються рідше, ніж очікувалося за незалежним успадкуванням. Він пов’язав це з фізичним обміном ділянками між гомологічними хромосомами. У 1913 році Стёртевант уперше побудував генетичну карту, розташувавши гени yellow, white та miniature відповідно до частот рекомбінації. Фізичне підтвердження обміну матеріалом надійшло в 1931 році: Барбара Мак-Клінток і Харріет Крейтон на кукурудзі, а Курт Штерн — на дрозофілі, показали, що рекомбінантні нащадки мають видимі структурні зміни в хромосомах. У 1933 році Морган отримав Нобелівську премію за ці відкриття, які лягли в основу хромосомної теорії спадковості.
Молекулярний механізм: від розриву ДНК до хіазм
Сучасне розуміння процесу спирається на модель подвійного розриву з репарацією (DSBR). Усе починається з того, що білок Spo11 спеціально нарізає обидва ланцюги ДНК у певних ділянках — гарячих точках рекомбінації. Ці розриви не випадкові: вони виникають у промоторах і GC-збагачених областях, де частота подій у тисячі разів вища, ніж у середньому по геному.
Комплекс MRN (або MRX у дріжджів) обробляє кінці розриву, видаляючи 5’-кінці та створюючи довгі 3’-одноланцюгові хвости. На ці хвости сідають рекомбінази Rad51 і мейоз-специфічна Dmc1, формуючи нуклеопротеїнові філаменти. Одна з ниток вторгається в гомологічну хромосому, витісняє комплементарний ланцюг і утворює D-петлю. Після синтезу ДНК і лігування виникають подвійні структури Холлідея (dHJ) — чотириниткові перехрестя.
У мейозі розв’язання цих структур відбувається переважно шляхом кросинговеру завдяки дії ендонуклеаз MLH1–MLH3 (шлях класу I) та MUS81 (шлях класу II). Синаптонемальний комплекс захищає проміжні структури від передчасного розв’язання, гарантуючи, що хоча б один кросинговер на бівалент відбудеться. Додатковий рівень регуляції забезпечує інтерференція: виникнення одного кросинговеру пригнічує появу іншого поблизу, розподіляючи події рівномірно вздовж хромосоми.
Цей механізм настільки точний, що навіть у великих геномах людини на кожну пару гомологів припадає в середньому один-два обов’язкові кросинговери, що запобігає катастрофічним помилкам сегрегації.
Роль кросинговеру в забезпеченні правильного поділу хромосом
Кросинговер виконує дві взаємопов’язані функції. По-перше, він генерує рекомбінантні гамети з новими комбінаціями алелів. По-друге — і це часто недооцінюють — хіазми фізично утримують гомологічні хромосоми разом до анафази I. Без цього «якоря» біваленти можуть розпастися передчасно, і хромосоми розійдуться випадково, спричиняючи анеуплоїдію — неправильну кількість хромосом у гаметах.
У багатьох видів, включно з людиною, існує механізм «обов’язкового кросинговеру»: кожна пара гомологів отримує щонайменше одну хіазму. Порушення цього правила — наприклад, через мутації в генах MSH4 або MSH5 — призводить до безпліддя, оскільки гамети формуються з помилками або гинуть. Таким чином кросинговер є не лише джерелом різноманітності, а й гарантом стабільності геному під час редукційного поділу.
Генетична рекомбінація та створення карт генів
Частота кросинговеру між двома генами прямо пропорційна відстані між ними на хромосомі. Це відкриття Стёртеванта лягло в основу генетичного картування. Одиницею виміру стала сантиморган (сМ) — відстань, на якій рекомбінація відбувається з імовірністю 1 %.
Однак реальна картина складніша через подвійні та множинні кросинговери, а також інтерференцію. Якщо два кросинговери відбуваються між одними й тими самими генами, фенотипічно це виглядає як відсутність рекомбінації. Тому для точних карт використовують триточкові схрещування та молекулярні маркери — SNP, мікросателіти. Сучасні геномні карти людини та модельних організмів побудовані саме на основі мільйонів зареєстрованих рекомбінаційних подій.
| Стадія профази I | Основні події | Роль кросинговеру |
|---|---|---|
| Лептотена | Конденсація хромосом, початок утворення розривів ДНК | Підготовка місць для майбутнього обміну |
| Зиготена | Кон’югація гомологів, формування синаптонемального комплексу | Створення платформи для точного спарювання хроматид |
| Пахітена | Повне спарювання, завершення кросинговеру | Безпосередній обмін ділянками між несестринськими хроматидами |
| Диплотена | Часткове роз’єднання хромосом, поява хіазм | Видимі наслідки обміну, утримання бівалентів |
| Діакінез | Максимальна конденсація, руйнування ядерної оболонки | Хіазми готові забезпечити правильну сегрегацію в метафазі I |
(Дані узагальнено з енциклопедичних джерел та публікацій у журналі Nature за 2024–2025 роки.)
Еволюційне значення та сучасні дослідження
Кросинговер — один із найпотужніших двигунів комбінативної мінливості. Він дозволяє «перетасовувати» алелі, зібрані протягом попередніх поколінь, створюючи нові генотипи за лічені покоління. Це особливо важливо для швидко змінюваних умов — паразитів, клімату, нових патогенів.
Паралельно кросинговер тісно пов’язаний з механізмами репарації ДНК: багато білків (Rad51, MLH-комплекси) працюють і в мітозі, і в мейозі. Еволюційно мейоз, ймовірно, виник як удосконалена система відновлення пошкоджень ДНК, яка водночас забезпечувала генетичну різноманітність.
У 2025 році в журналі Nature опублікували дослідження, яке показало, як синаптонемальний комплекс і когезини захищають подвійні структури Холлідея від передчасного розв’язання не на користь кросинговеру. Це пояснює, чому навіть за наявності альтернативних шляхів (SDSA, розчинення) більшість подій у мейозі завершуються саме обміном.
Дослідження на людях виявили, що варіанти генів, які контролюють гарячі точки рекомбінації (зокрема PRDM9), впливають на фертильність і ризик хромосомних аномалій. У селекції рослин і тварин знання про кросинговер дозволяє цілеспрямовано розривати небажані зчеплення генів і прискорювати створення нових сортів та порід.
Цікаві факти про кросинговер
- У самців плодової мушки Drosophila melanogaster кросинговер повністю відсутній — унікальний випадок, коли рекомбінація залежить від статі. Самки ж демонструють типову частоту обміну.
- Фізичне підтвердження обміну генетичним матеріалом уперше отримали в 1931 році на кукурудзі та дрозофілі: рекомбінантні нащадки мали видимі цитологічні зміни в структурі хромосом.
- Кожна пара гомологічних хромосом у більшості еукаріотів отримує щонайменше одну обов’язкову хіазму — це механізм гарантії правильного розходження, без якого ризик анеуплоїдії різко зростає.
- Гарячі точки рекомбінації займають менше 1 % геному, але саме там відбувається переважна більшість кросинговерів; їх розташування частково контролюється білком PRDM9 у ссавців.
- Мутації в генах MSH4 та MSH5, що стабілізують структури Холлідея, у людини призводять до безпліддя через масове порушення утворення гамет.
- У 2025 році дослідження в Nature показало, що синаптонемальний комплекс буквально «захищає» проміжні структури кросинговеру від конкурентних шляхів розв’язання, забезпечуючи надійність процесу.
- Іонізуюче випромінювання та певні хімічні речовини можуть підвищувати частоту кросинговеру, впливаючи на репарацію ДНК, — це явище використовують у експериментальній генетиці для збільшення варіабельності.
Кросинговер продовжує залишатися однією з найбільш елегантних і водночас складних систем природи — процесом, де точність молекулярних машин поєднується з творчою силою генетичної мозаїки, що лежить в основі всього розмаїття життя на Землі.